CONTENT
| The group "Georgians" in 1082 AD. (914—1278 AD.)
| A history of the residence region of the group "Georgians" during their main admixture event in 1082 AD. (914—1278 AD.)
| The group "Adygei" in 1530 AD. (1418—1810 AD.)
| A history of the residence region of the group "Adygei" during their main admixture event in 1530 AD. (1418—1810 AD.)
| The group "Adygei" in 990 BC. (3678 BC. — 830 AD.)
| A history of the residence region of the group "Adygei" during their main admixture event in 990 BC. (3678 BC. — 830 AD.)
| Admixture conclusions on "Adygei" and "Georgian" groups
Y chromosome
| Genetic pictures of the Abkhaz-adyghe and the south caucasian populations
| The Abkhazian Y-chromosome haplogroups
| Time to the common male ancestor by 111 markers on G-M201 for Abkhazians and Georgians
| Western Caucasian (Circassians, Abkhazians, Megrelians, Georgians, Ossetians) Y-chromosome haplogroups conclusions
Mitochondrial DNA
| Abkhazian mitochondrial DNA haplogroups
| Abkhazian mitochondrial DNA haplogroups conclusions
History and languages
| History and languages conclusions
General conclusions
Materials
The Abkhaz and Western Caucasian DNA
«Facts do not cease to exist because
they are ignored» — Aldous Huxley
«He who controls the past
controls the future.» — George Orwell
they are ignored» — Aldous Huxley
«He who controls the past
controls the future.» — George Orwell
One of the most difficult issues of the modern history is a determination of a degree of the caucasian populations kinship. This anthropogenetical study will try to make one more step in solving this problem, relying on a broad base of genetic researches, especially of male Y-chromosome markers, which are one of the most informative markers in the reconstruction of population kinships.
Admixtures
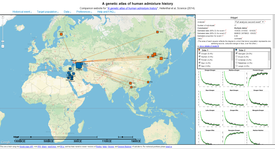
14 February 2014, the "Science" journal has published this article with a link to A genetic atlas of human admixture history. The job was carried out under the supervision of geneticist Dr. Garrett Hellenthal and is described in this document and a FAQ. They used data analysis autosomal single nucleotide polymorphism (autosomal SNPs) from 1490 people out of 95 population groups (2 to 46 people in each group). They define the group as "a certain number of people with an identical genetic picture" and date back the main genes admixing events between the studied groups.
- Example 1
www.admixturemap.paintmychromosomes.com / Menu / Historical event / Tutorial part 1 - simple events
"...For example, look at the 'Moroccan-Yoruba' curve in the box on the left (Example 1). The upward slope of this curve indicates that Moroccan-like and Yoruban-like DNA were on opposite "sides" of the event. Similarly, the Moroccan-Tunisian curve slopes downwards implying that these segments of DNA came from the same source..."
In other words, the orange objects are always on one side of the main admixing events, and the blue are on the opposite. These data, in combination with the historical materials, will show a more accurate picture on the ethnogenesis of those groups in the given periods of their main admixing events.
A short dialog with Dr. Hellenthal, clarifies some more details of their data analyzing methods:
On Wednesday, May 21, 2014, Давид Аршба <darshba@XXX> wrote:
Good afternoon, Dr. Hellenthal.
I am intrested in two groups within your project (www.admixturemap.paintmychromosomes.com): Adygei and Georgian. Georgian has Estimated date (95% CI): 1082CE (914CE - 1278CE). Initially I was thinking that this means that Georgian group was formed out of all inferred groups in the timeline 914CE - 1278CE, and the 1082CE is an average date on this timeline. But now I've found out that this might be the only one admixture event, which was stretched on 914CE - 1278CE, and the 1082CE date is not an average date. Could you briefly decribe the method of finding this date? And a second question: why did you choose to analyze precisely this period of admixture event and not the other? As I understand, those admixture events can occur anytime a new child is born.
Thank you for such world overturning researches. Good luck!
David Arshba
---
On Wednesday, May 28, 2014, Garrett Hellenthal <ghellenthal@gmail.com> wrote:
Hi David -
Thanks for your interest!
For your date question, our method infer specific admixture events. These are when two (or perhaps more) genetically different groups intermix and have children. Our model assumes this happens at a precise time (i.e. a particular generation, though the model can be more flexible) in the past. In reality if two different groups meet, in some cases it could take some years for them to intermix at a level our approach can detect. In such a case, the date we infer likely represents a time slightly after (i.e. more recent than when) they first met up and started mixing.
For our Georgian samples, we infer that our assumption that intermixing happens over a single generation - though clearly wrong - fits the data quite well and so likely isn't too far from the truth. Our "best-guess" for the date of this mixing event is 1082CE. However, there is statistical uncertainty in our date estimate, such that the true date of this intermixing (likely) falls somewhere within 914-1278CE. Again this date range is meant to reflect a single event where two groups met and exchanged DNA over a narrow time frame (i.e. over tens of years). There may be other older events in our Georgian data that we miss, as our approach has more power to see recent intermixing events. But at least with our sample size of 20 individuals, we do not see evidence for any other (older) mixing events.
In contrast, for the Adygei we DO see evidence for two distinct mixing events, one occuring at (or around) 1530CE and the other much older (~990BCE). However, if you could see the SOM, it would caution that in this case because we infer that similar groups intermixed at each of these two times, it is possible that -- instead if two distinct events - there has been continuous mixing from East Asia into this region from ~990BCE until ~1530CE. So interpreting these signals can be complicated!
hope this helps,
Garrett
The group "Georgians" in 1082 AD. (914—1278 AD.)
- Graph 1
Number of individuals: 20
The time period of the admixing events: 914—1278 AD.
Approximate date of the main admixing event: 1082 AD.
The group "Georgians", as a modern population was formed as a result of the main admixing event in 1082 AD. between 914 AD. and 1082 AD. from the following groups:
The first side of the admixtures of the "Georgians" (orange): Adygei (28.6%), Palestinian (0.4%), Hazara (0.4%), Brahui (0.3%), Naxi (0.3%), Druze (0.3%), IndianJew (0.1%), Indian (0.1%), Yi (0.1%), Hezhen (0.1%)
The second side of the admixtures of the "Georgians" (blue): Greek (19.5%), Armenian (12.4%), SouthItalian (6.3%), Cypriot (5.3%), Lezgin (5.1%), NorthItalian (3.9%), Syrian (3.0%), Jordanian (2.6%), EastSicilian (2.4%), Pathan (1.7%), Makrani (1.4%), Tuscan (1.3%), Druze (0.9%), Sindhi (0.8%), Bedouin (0.7%), Burusho (0.6%), Palestinian (0.3%), Brahui (0.2%), Kalash (0.2%), Balochi (0.2%), WestSicilian (0.2%)
The region history of the "Georgians" residence during the main admixture event in 1082 AD. (914—1278 AD.)
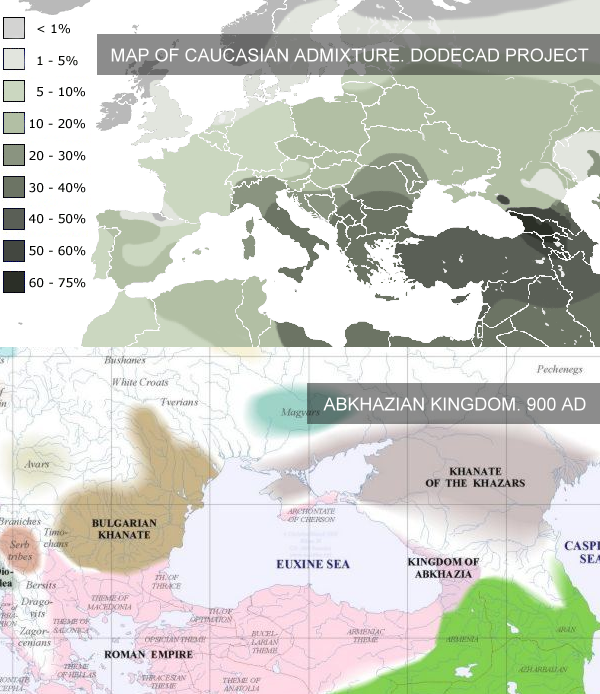
According to Dr. Hellenthal, from the dialog above (... For our Georgian samples ... at least with our sample size of 20 individuals, we do not see evidence for any other (older) mixing events ...) This admixing event of the group "Georgians" exactly coincides with the period of the Abkhazian Kingdom and it's later evolve to the Kingdom of Abkhazians and Kartvelians (Sakartvelo) which was located almost on 80% of today's Georgia's territory, included Abkhazia and part of Turkey to Trabzon (See additional materials below. Achugba, Bakhtadze, Vachnadze, Guruli). The Graph 1 shows the three main parts of a admixture of the "Georgians" group. Those groups are Adygei, Armenians and Greeks.
- Example 2
The dominant segments of the "Adygei" and the "Armenians" can be explained by the fact that during this period the region of Central Georgia was constantly passing from Armenian to Abkhazian kings and vice versa. The presence of "Greeks" in the region, is also quite evident throughout the studied period. Those kings where partly Greeks themselves as well as their armies, religious leaders and of course the Pontic Greeks living in the region since the 5th century BC.
The group "Adygei" in 1530 AD. (1418—1810 AD.)
- Graph 2

Number of individuals: 17
The time period of the admixing events: 1418—1810 AD.
Approximate date of the main admixing event: 1530 AD.
The group "Adygei", as a modern population was formed as a result of two main admixing events. The second more ancient event, described further below. The first event in 1530 AD. between 1418 AD. and 1810 AD. and the admixing groups were:
The first side of the admixtures of the "Adygei" (orange): Finnish (1.8%), Xibo (1.5%), Oroqen (1.4%), Mongola (0.9%), Hazara (0.6%), Yi (0.5%), Yakut (0.2%), Balochi (0.2%), Hezhen (0.2%), Lithuanian (0.2%), Orcadian (0.2%), Burusho (0.2%), Basque (0.1%), Daur (0.1%), Maya (0.1%), BantuKenya (0.1%), Indian (0.1%), Lahu (0.1%), Chuvash (0.1%), Colombian (0.1%), Hadza (0.1%), Naxi (0.1%)
The second side of the admixtures of the "Adygei" (blue): Georgian (35.8%), Hungarian (13.3%), Turkish (11.8%), Armenian (5.9%), Lezgin (5.8%), Greek (5.6%), Cypriot (3.0%), Iranian (2.2%), Pathan (1.3%), Balochi (1.3%), Syrian (1.1%), Polish (0.9%), Chuvash (0.8%), Palestinian (0.5%), Scottish (0.5%), Druze (0.4%), Sindhi (0.3%), Burusho (0.3%), Kalash (0.2%)
The region history of the "Adygei" residence during the main admixture event in 1530 AD. (1418—1810 AD.)
Graph 2 shows the 3 main components of the admixture of the "Adygei" for the mentioned period: Georgians, Hungarians and Turks. The similarity of the genetic picture with the group "Georgians" is clarified in the previous section. That means, if the group "Georgians" was formed as the modern population in the periods of the cooperative Abkhaz-Georgian kingdoms (914—1278 AD.), then the later period (1418—1810 AD.), logicaly shows the predominance of the group "Georgians" in today's group "Adygei".
The turkic segment is associated with the long time common history. It is known from the ancient history of Abkhazia that the Abkhazian king Leon I had married the daughter of the Khazar Kagan (VIII cent.). Their nephew Leon II with the support of the khazars getting the secession of Abkhazia and Lazica from the Byzantine Empire (VIII cent.) and declares independence of the Abkhazian kingdom. The more close contacts of the Abkhaz and the Turks dates back to XV century in the period of Ottoman Empire (XVI—XVIII centuries.). According to A.K. Shagirov most of the turkisms got to the Adygei during the Mongol-Tatar and the Crimean-Turkish conquests of the Northwest Caucasus [Shagirov 1989, p. 74]. During XIII-XIV centuries the North Caucasus is being settled by turkic-speaking Balkars, Karachais and Polovtsians, which are associated with modern Kumyks. In about XIV-XV centuries the Nogai appear here as well. Later on the turkic-speaking Truhmens arrive to Stavropol, the relatives of central asian Turkmens [Klimov 1965, p. 11]. King David IV the Builder in the fight against the Oghuz Turks of the Seljuk Empire invited and gave a permission to settle in Borchali and adjacent areas the Kipchak horde numbering 40 000 soldiers, which in total counting their families would be about 200 000 people. It is appropriate to note that at that time King David IV had also married the daughter of Kipchaks khan.
The segment of the group "Hungarians". From the 9th century the Magyars (proto Hungarians) migrated to the Circassian steppes of Northern Black Sea region. In 9-10 centuries they roamed the steppes between the Don and the Dnieper and the Khazars in 835 AD. built the Sarkel fortress on the left bank of the Don (Tsymlyanskoe) to protect themselves from Magyars. It is also known that in 1236 AD. Hungarian dominicans (catholic monastic order) went to the Volga river and started their missions in Matreg (or Tmutarakan - one of the oldest cities in the Taman Peninsula, which was located in present-day Taman Temryuk district of Krasnodar region). Also the Matreg Catholic Diocese with the a Circassian bishop was formed in the Taman peninsula. The part of the modern Circassians, for example, Besleneys (adygei subethnos), is generally considered to be descendants of the Magyars.
The group "Adygei" in 990 BC. (3678 BC. — 830 AD.)
- Graph 3
Number of individuals: 17
The time period of the admixing events: 3678 BC. — 830 AD.
Approximate date of the main admixing event: 990 BC.
The group "Adygei", as a modern population was formed as a result of two main admixing events. The first event is described above. The second event in 1530 AD. between 1418 AD. and 1810 AD. and the admixing groups were:
The first side of the admixtures of the "Adygei" (orange): Oroqen (6.4%), Hezhen (5.5%), Finnish (4.2%), Daur (4.2%), Naxi (3.9%), Yakut (3.3%), Lahu (2.7%), She (2.7%), Mongola (2.6%), Xibo (2.4%)
The second side of the admixtures of the "Adygei" (blue): Georgian (24.0%), Druze (5.2%), Armenian (5.2%), Palestinian (4.3%), Lezgin (4.2%), Turkish (4.2%), Syrian (3.9%), Bedouin (3.7%), Cypriot (3.7%), Jordanian (3.6%)
The region history of the "Adygei" residence during the main admixture event in 990 BC. (3678 BC. — 830 AD.)
Graph 3 shows the group "Georgians" as the main component of the admixture of the "Adygei" in 990 BC. (3678 BC. — 830 AD.). Its presence and the similarity with the modern group "Georgians" was discussed in the two previous analysis of admixtures. The other segments (76%), due to the very large time period between 3678 BC. and 830 AD. are present in the admixture with approximately equal amounts.
Admixture conclusions on "Adygei" and "Georgian" groups
The names of the groups (the Georgians, Adygei, etc.) are defined by researchers of "A genetic atlas of human admixture history" in their FAQ in the section "How have you defined a “group”?" They define the group as "a certain number of people with an identical genetic picture" and date back the main genes admixing events between the studied groups.
1. There is a same main segment of admixtures in the studied groups "Adygei" and "Georgians". It is called "Adygei" in the group "Georgians" and "Georgians" in the group "Adygei".
2. The group "Georgians", as a modern population was formed as a result of the main admixing event in 1082 AD. between 914 AD. and 1082 AD.
3. According to Dr. Hellenthal, they haven't find the evidence for any other (older) mixing events in the group "Georgians" than 1082 AD. (914—1278 AD.).
4. Thу main admixing event of the group "Georgians" exactly coincides with the period of the Abkhazian Kingdom and it's later evolve to the Kingdom of Abkhazians and Kartvelians (Sakartvelo).
5. The group "Adygei", as a modern population was formed as a result of two main admixing events: 1530 AD. (1418—1810 AD.) and more ancient event 990 BC. (3678 BC. — 830 AD.).
6. The presence in the "Adygei" admixture of the older cases of admixing events compared with the group "Georgians" suggests that today's Georgians (formed as a result of the main population admixing event in 1082 for the period from 914 till 1278) are mainly Adygei genetically, but with a modern accumulated differences in cultural and linguistic forms.
7. The identical predominant type of admixture in both groups can mean the following:
- Option 1 - in 1082 AD. (91—1278 AD.) in the period of Abkhazian Kingdom the Georgian population was gradually absorbed by the Abkhaz-Adyghe, which led to the modern genetic picture of Georgians.
- Option 2 - before the above mentioned period (91—1278 AD.) the groups "Georgians" and "Adygei" has already been constituted as a unified nation.
8. The differences of cultures and languages on the background of an identical type of admixture in both groups, is likely explained by the political decisions of those days rulers.
9. About 75% of the remaining admixture segments from both groups, are presumably being the cause of differences in anthropological types between these them.
10. The full analysis of the main types of admixtures defines those groups as Near Eastern or Mesopotamian.
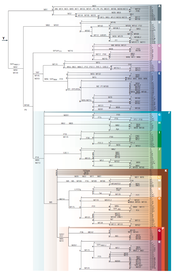
- Y-DNA Phylogenetic Tree
Y chromosome
Brief definitions: autosomal chromosomes, mitochondrial DNA (mtDNA), X chromosomes, Y chromosomes, mutations, Y-DNA haplogroups by ethnic groups and countries, Caucasian Y-DNA haplogroups
Women and men have 23 pairs of chromosomes, where one chromosome was received from the father and the other from the mother. Unlike autosomal chromosomes, that are called by numbers from "1" to "22", the two of "sex" chromosomes have letter designations. XX for females and XY for males. The child inherits from the father the X chromosome (girl) or the Y chromosome (boy). The X chromosome from the father turns into a combination of XX and gives a birth to a girl. The Y chromosome from the father turns into a combination of XY and gives a birth to a boy. Almost all chromosomes undergo mixing (recombination) - it is a process when each pair of chromosomes exchange their fragments with each other. Since every male has only one Y chromosome it does not recombine, in contrast to the X chromosomes. For these reasons, a genealogical analysis using the X chromosomes is much more complicated. We also inherit mitochondrial DNA (mtDNA) from the mother, and none of them from the father.
The main tools of DNA genealogy tests are the mutations, their number and location in the mtDNA and
- Y-DNA Migration Map
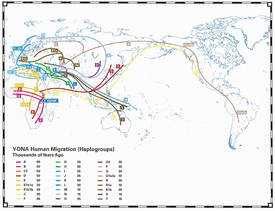
An additional opportunity for such studies may be the ancient archaeological finds from preserved DNA, such as Ötzi - the ice mummy man Chalcolithic era,
discovered in 1991 in the Tyrolean Alps. Ötzi belonged to haplogroup G2a and his age, found by radiocarbon method, is about 5300 years old. There are also finds of the same period, but in Spain (Lacan 2011, Haak 2010), where in the complex of burials they found bodies with the same G2a haplogroup. Also, recently in Bavaria were found 6 buried heavy riders cataphracts. 4 of 6 belonged to haplogroup R1b, the other two were of G2a (the main for the Western Caucasus) alongside whom the burial of horses, that were sacrificed in the sarmatian style (head and legs neatly stacked). (Vanek 2009)
As for today the Abkhazians, as well as the West Caucasus have mainly G2a [50,6] in their samples and three other haplogroups: J2 [22,3], R1a1a [9,9], R1b1b2 [6,4]. The highest percentage of representatives of haplogroup G2a common among Shapsugs (about 81%), and among the Abkhazians and Ossetians (about 55-75%).
By Litvinov 2010 (see Materials below):
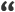
Haplogroup G-M201 is divided into two major branches with a distinctive geography: G2-P287 and G1-M285, defined with M285 and M342 mutations [Karafet et al., 2008]. Haplogroup G1-M285 of the populations of Western Caucasus is found only in rare cases in populations of Adygei and Kabardians [Bogotova, 2009], and nowhere else recorded, but reaching 10.5% of Armenians (6 out of 57 individuals). Given that the population we studied is rather limited in numbers, it needs more precise interpretation of such a high frequency, taking into account a possible founder effect. Regarding the G2-P287 haplogroup in the Western Caucasus, it was presented by G2a-P15 and G2 * -P287 (xG2a) haplogroups. Comparing dissemination of those subclades the attention is needed to the fact that, despite an overall widespread of the haplogroup G2a-P15 and G2 * -P287 (xG2a), Abkhaz-Adygei characterized by a high incidence of G2 *-P287 (xG2a), and the other nations of the Western Caucasus are mostly G2a-P15. While here it is important to note that the G2 * -P287 (xG2a) with a frequency of 10.3% seen in the Kuban Nogai, but considering their Central Asian origin, itis most likely a later inclusion of this haplogroup in the genetic pool of the Nogai, since they were in close contact with the autochthonous Caucasian population. However, without knowing the phylogenetic status of haplogroup G2 * -P287 (xG2a), we can not make any conclusion of its origin. It is very likely, considering the results of [url = http: //arshba.ru/y-chromosome-haplotype-strata-in-anatolia-cinnioglu-2003-t470.html]. Cinnioglu et al [/ url], that G2a-P15 reflects the spread of Hattians culture in West Caucasus, all the more that this haplogroup is rare in Dagestan, reaching only at Andi and Lezghins 6.1 and 9.7% respectively, while the rest of Dagestan frequency of G2a-P15 does not exceed 5 %Yunusbaev, 2006; Kutuev et al., 2010]. On the other hand, there are not many populations worldwide with the presence of haplogroup G2 * -P287 (xG2a), this may indicate that there is a certain subclade inside the G2-P287, specific only to the Western Caucasus and, presumably, was formed on the basis of the Abkhaz-Adyghe substrate.
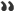
- Fig. 5. The gene pool of Abkhaz-Adyghe populations, Georgians and Armenians
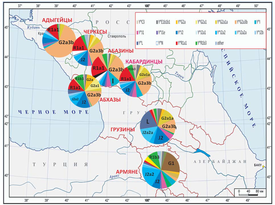
Genetic portraits of Abkhaz-Adyghe and Transcaucasia populations
By Теучеж 2013:
Haplogroup G. An analysis of the Y-chromosome variation found 35 haplogroups. "Genetic Portraits" of studied populations - the main range of the identified haplogroups and their shares in the gene pool are shown in Fig. 5. A significant part of the gene pool of the Abkhaz-Adyghe populations represented by a western asian haplogroup G2a3b-P303, which is consistent with previous results of our studies [Dibirova et al., 2010; Balanovsky et al., 2011]. In this case the maximum frequency detected in the three sub-ethnic groups of Adygei, in which it varies from 53% to 86%. Kabardians, Circassians, Abkhaz and Abaza populations have lower frequency of G2a3b-P303 - from 18% to 30%. Among other peoples of the Caucasus, this haplogroup is virtually absent: the peoples of Dagestan Avars have 10%, in Chechnya and Ingushetia, its frequency is an average of 2%, Ossetians - 3%. However, in the gene pool of Balkar the haplogroup frequency is increased to 17% [Shalyaho et al., 2013], ie almost to the same level as that of the neighboring Kabardians (22%). Georgians frequency of the haplogroup G2a3b-P303 (13%) is only slightly lower than that of the Abkhazians (18%), and the Armenians, it is practically absent. Armenians have another big branch of haplogroup G - haplogroup G1-M285 (20%).
- Table. 1. Genetic distances of 35 Western Caucasus Y-chromosome haplogroups
Haplogroup J. Another Near Eastern haplogroup found in the Abkhaz-Adyghe populations is J2. It was found with an average of 15%. However, in the Abkhazians it reaches a high frequency (27%), which brings them together with the Georgians and Armenians, that also have this haplogroup in about one third of their gene pool. The maximum values in the Caucasus frequency of this haplogroup reaches among the population of the Eastern Caucasus - Ingush (89%) and Chechens (55%).
A feature of the gene pool of the Abkhazians is the increase of the haplogroup J2a2-M67 (12%), that in the Armenians have 20%, reaching a maximum frequency in the Western Armenian Hamshen (31%). This haplogroup is virtually absent in Georgian populations we studied: they have a different version of this haplogroup J2a2a-M92 (13%).
Western Asian haplogroup J1-M267: Kubachins (98%) [Balanovsky et al., 2011]), Chechens (98%), Abkhaz-Adyghe (0-5 %).
- Fig. 6. Comparison of the populations of Asia Minor, Europe and the Caucasus on 14 Y-chromosome haplogroups
Haplogroup R1a. A significant contribution to the gene pool of the Abkhaz-Adyghe found by the Western Eurasian haplogroup R1a1-M198, averaging 15% of the total gene pool, and reaching 25% in Abaza. There are 33% samples of this haplogroup among the Turkic peoples of the Western Caucasus [Схаляхо с соавт., 2013]. The presence of this haplogroup, according to some authors, may reflect the influence of Iranian-speaking population of the Eurasian steppes, preceding the expansion of the Turkic and Mongolian peoples [Багашев, 1998; Харьков с соавт., 2007]. A modest contribution of this haplogroup in the gene pool of the peoples of Transcaucasia (2-4%) may indicate the validity of this hypothesis. R1a1a1g-M458, which is the main european branch of haplogroup R1a1a-M198 [Underhill et al., 2009] and the half of the Slavs gene pool, was found in around 2-5% among Adygei, Kabardians, Circassians, Abkhazians, Abaza, Georgians and the Armenians 0%. In the Caucasus, haplogroup R1a1a1g-M458 reaches a high level only in the Kuban Nogais - 18%. The gene pool of the remaining populations of this haplogroup is found at low frequencies from 1% in Kumyks to 6% in the Balkars [Схаляхо c соавт., 2013]. This suggests that the gene pools of populations we have studied almost found no genetic traces of Slavic influences.
Other western eurasian haplogroups such as R1b3-M269, as well as a total of all subclades of a haplogroup I (including I1, I2a, I2b) are found from 0% to 9% (an average of 5%). Eastern eurasian haplogroup C3-M217 and C3C-M48 are only 0.5% in the studied populations. The average frequency of northern eurasian haplogroups Q-M242 and N1C1-M178 is 1%. With the general similarity of the distribution of haplogroups there are also some differences between populations. The greatest variety of haplogroups among Abkhaz-Adyghe populations are found in Kabardians and Circassians (28 Y-chromosome haplogroups). However, only the Kabardians have an unusual variety of caucasian haplogroups O3, G1a and F.
The Abkhazian Y-chromosome haplogroups
Comparison of Y-DNA chromosomes of Abkhazians, Circassians, Mingrelians, Georgians and Ossetians
The Time to a most recent common ancestor (TMRCA) for 111 markers of G-M201 samples for Abkhazians and Georgians
Distances are determined by using these scripts ysearch, isogg using the methods described in Walsh 2000 and Zhivotovsky 2004. The main data is from Balanovsky 2011 (Supplementary_Tables.xls), FTDNA Georgian DNA Project, FTDNA Ossetian DNA Project, FTDNA Adyghe DNA Project
In the following calculations of time to a most recent common ancestor - Cerkesoglu, Karba, Abashidze, K Shapsug and Khintba will be used from the Abkhaz side for comparison with the georgian side Swann, Ossetians, Mingrelians, Laz people and other G-M201 populations from Georgia.
Table 1
Kit Number |
Paternal Ancestor Name |
Country |
Haplogroup |
DYS393 |
DYS390 |
DYS19 |
DYS391 |
DYS385 |
DYS426 |
DYS388 |
DYS439 |
DYS389i |
DYS392 |
DYS389ii |
DYS458 |
DYS459 |
DYS455 |
DYS454 |
DYS447 |
DYS437 |
DYS448 |
DYS449 |
DYS464 |
DYS460 |
Y-GATA-H4 |
YCAII |
DYS456 |
DYS607 |
DYS576 |
DYS570 |
CDY |
DYS442 |
DYS438 |
DYS531 |
DYS578 |
DYF395S1 |
DYS590 |
DYS537 |
DYS641 |
DYS472 |
DYF406S1 |
DYS511 |
DYS425 |
DYS413 |
DYS557 |
DYS594 |
DYS436 |
DYS490 |
DYS534 |
DYS450 |
DYS444 |
DYS481 |
DYS520 |
DYS446 |
DYS617 |
DYS568 |
DYS487 |
DYS572 |
DYS640 |
DYS492 |
DYS565 |
DYS710 |
DYS485 |
DYS632 |
DYS495 |
DYS540 |
DYS714 |
DYS716 |
DYS717 |
DYS505 |
DYS556 |
DYS549 |
DYS589 |
DYS522 |
DYS494 |
DYS533 |
DYS636 |
DYS575 |
DYS638 |
DYS462 |
DYS452 |
DYS445 |
Y-GATA-A10 |
DYS463 |
DYS441 |
Y-GGAAT-1B07 |
DYS525 |
DYS712 |
DYS593 |
DYS650 |
DYS532 |
DYS715 |
DYS504 |
DYS513 |
DYS561 |
DYS552 |
DYS726 |
DYS635 |
DYS587 |
DYS643 |
DYS497 |
DYS510 |
DYS434 |
DYS461 |
DYS435 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 163132 | Mustafa Cerkesoglu (Husht) Circassia Shapsug, Trabzon | Turkey | G-M201 | 14 | 23 | 15 | 10 | 13-14 | 11 | 12 | 11 | 13 | 11 | 31 | 17 | 9-9 | 11 | 11 | 23 | 16 | 20 | 29 | 13-13-14-14 | 11 | 11 | 20-20 | 16 | 13 | 16 | 20 | 35-36 | 11 | 10 | 11 | 8 | 15-16 | 8 | 12 | 10 | 8 | 12 | 11 | 14 | 21-22 | 17 | 10 | 12 | 13 | 14 | 8 | 12 | 21 | 24 | 17 | 13 | 11 | 14 | 10 | 11 | 11 | 11 | 35 | 15 | 8 | 17 | 12 | 21 | 27 | 21 | 11 | 10 | 12 | 13 | 11 | 9 | 9 | 11 | 10 | 10 | 12 | 27 | 10 | 12 | 21 | 14 | 11 | 13 | 29 | 15 | 18 | 16 | 24 | 19 | 13 | 15 | 27 | 12 | 22 | 18 | 11 | 14 | 17 | 9 | 11 | 11 |
| 306115 | Chaguna Karbaia, b. c. 1775, Darcheli, Samegrelo | Georgia | G-M201 | 14 | 23 | 15 | 10 | 13-13 | 11 | 12 | 11 | 12 | 11 | 28 | 18 | 9-9 | 11 | 11 | 23 | 16 | 20 | 31 | 13-13-13-14 | 10 | 11 | 20-21 | 17 | 12 | 15 | 18 | 35-36 | 12 | 10 | 11 | 8 | 15-16 | 8 | 11 | 10 | 8 | 12 | 11 | 14 | 21-22 | 15 | 10 | 12 | 12 | 13 | 8 | 12 | 21 | 23 | 19 | 13 | 11 | 13 | 10 | 11 | 11 | 12 | 29 | 15 | 8 | 16 | 12 | 21 | 27 | 21 | 11 | 11 | 11 | 13 | 11 | 9 | 9 | 11 | 10 | 10 | 12 | 27 | 10 | 11 | 21 | 14 | 11 | 13 | 26 | 15 | 19 | 14 | 24 | 18 | 13 | 15 | 27 | 12 | 21 | 18 | 11 | 14 | 16 | 9 | 12 | 11 |
| TMRCA - Time to Most Recent Common Ancestor / Расстояние до ближайшего общего предка (мутация 0.004..0.0075 FTDNA; поколение 24-30 лет) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1060 ± 212 лет | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 163132 | Mustafa Cerkesoglu (Husht) Circassia Shapsug, Trabzon | Turkey | G-M201 | 14 | 23 | 15 | 10 | 13-14 | 11 | 12 | 11 | 13 | 11 | 31 | 17 | 9-9 | 11 | 11 | 23 | 16 | 20 | 29 | 13-13-14-14 | 11 | 11 | 20-20 | 16 | 13 | 16 | 20 | 35-36 | 11 | 10 | 11 | 8 | 15-16 | 8 | 12 | 10 | 8 | 12 | 11 | 14 | 21-22 | 17 | 10 | 12 | 13 | 14 | 8 | 12 | 21 | 24 | 17 | 13 | 11 | 14 | 10 | 11 | 11 | 11 | 35 | 15 | 8 | 17 | 12 | 21 | 27 | 21 | 11 | 10 | 12 | 13 | 11 | 9 | 9 | 11 | 10 | 10 | 12 | 27 | 10 | 12 | 21 | 14 | 11 | 13 | 29 | 15 | 18 | 16 | 24 | 19 | 13 | 15 | 27 | 12 | 22 | 18 | 11 | 14 | 17 | 9 | 11 | 11 |
| 299046 | Khuta Gulordava, b. c. 1770, Kitsia, Samegrelo | Georgia | G-M201 | 14 | 22 | 15 | 10 | 16-16 | 11 | 12 | 11 | 13 | 10 | 30 | 17 | 9-9 | 11 | 12 | 24 | 16 | 21 | 28 | 13-13-14-14 | 10 | 10 | 20-21 | 16 | 15 | 15 | 18 | 36-37 | 11 | 10 | 11 | 8 | 15-16 | 8 | 11 | 11 | 8 | 11 | 10 | 12 | 20-21 | 15 | 10 | 12 | 12 | 16 | 8 | 13 | 21 | 22 | 16 | 13 | 11 | 13 | 10 | 11 | 11 | 13 | 30 | 16 | 8 | 16 | 11 | 23 | 28 | 20 | 9 | 11 | 13 | 12 | 12 | 9 | 10 | 11 | 10 | 11 | 12 | 28 | 10 | 12 | 22 | 14 | 9 | 10 | 24 | 15 | 17 | 12 | 23 | 18 | 12 | 16 | 27 | 12 | 21 | 18 | 11 | 14 | 17 | 9 | 12 | 11 |
| TMRCA - Time to Most Recent Common Ancestor / Расстояние до ближайшего общего предка (мутация 0.004..0.0075 FTDNA; поколение 24-30 лет) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2568 ± 642 года | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 299046 | Khuta Gulordava, b. c. 1770, Kitsia, Samegrelo | Georgia | G-M201 | 14 | 22 | 15 | 10 | 16-16 | 11 | 12 | 11 | 13 | 10 | 30 | 17 | 9-9 | 11 | 12 | 24 | 16 | 21 | 28 | 13-13-14-14 | 10 | 10 | 20-21 | 16 | 15 | 15 | 18 | 36-37 | 11 | 10 | 11 | 8 | 15-16 | 8 | 11 | 11 | 8 | 11 | 10 | 12 | 20-21 | 15 | 10 | 12 | 12 | 16 | 8 | 13 | 21 | 22 | 16 | 13 | 11 | 13 | 10 | 11 | 11 | 13 | 30 | 16 | 8 | 16 | 11 | 23 | 28 | 20 | 9 | 11 | 13 | 12 | 12 | 9 | 10 | 11 | 10 | 11 | 12 | 28 | 10 | 12 | 22 | 14 | 9 | 10 | 24 | 15 | 17 | 12 | 23 | 18 | 12 | 16 | 27 | 12 | 21 | 18 | 11 | 14 | 17 | 9 | 12 | 11 |
| 306115 | Chaguna Karbaia, b. c. 1775, Darcheli, Samegrelo | Georgia | G-M201 | 14 | 23 | 15 | 10 | 13-13 | 11 | 12 | 11 | 12 | 11 | 28 | 18 | 9-9 | 11 | 11 | 23 | 16 | 20 | 31 | 13-13-13-14 | 10 | 11 | 20-21 | 17 | 12 | 15 | 18 | 35-36 | 12 | 10 | 11 | 8 | 15-16 | 8 | 11 | 10 | 8 | 12 | 11 | 14 | 21-22 | 15 | 10 | 12 | 12 | 13 | 8 | 12 | 21 | 23 | 19 | 13 | 11 | 13 | 10 | 11 | 11 | 12 | 29 | 15 | 8 | 16 | 12 | 21 | 27 | 21 | 11 | 11 | 11 | 13 | 11 | 9 | 9 | 11 | 10 | 10 | 12 | 27 | 10 | 11 | 21 | 14 | 11 | 13 | 26 | 15 | 19 | 14 | 24 | 18 | 13 | 15 | 27 | 12 | 21 | 18 | 11 | 14 | 16 | 9 | 12 | 11 |
| TMRCA - Time to Most Recent Common Ancestor / Расстояние до ближайшего общего предка (мутация 0.004..0.0075 FTDNA; поколение 24-30 лет) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2208 ± 552 года | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 216684 | Kaikhosro Abashidze, b.c. 17??, Georgia | Georgia | G-M201 | 13 | 23 | 15 | 10 | 15-16 | 11 | 12 | 10 | 12 | 10 | 28 | 18 | 9-9 | 11 | 11 | 24 | 16 | 21 | 29 | 13-14-14-14 | 10 | 10 | 21-21 | 14 | 15 | 14 | 18 | 34-37 | 12 | 10 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 283050 | Dzikva Khintba, Abkhazia, Okhurei | Abkhazia | G-M201 | 14 | 24 | 15 | 10 | 12-16 | 11 | 12 | 11 | 12 | 12 | 29 | 17 | 9-9 | 11 | 11 | 23 | 16 | 20 | 29 | 13-13-13-13 | 10 | 11 | 20-21 | 15 | 13 | 16 | 19 | 34-37 | 11 | 10 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TMRCA - Time to Most Recent Common Ancestor / Расстояние до ближайшего общего предка (мутация 0.004..0.0075 FTDNA; поколение 24-30 лет) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1656 ± 414 лет | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 283822 | Almaskhan Buzaladze Born 1870 | Georgia | G-P303 | 14 | 22 | 15 | 11 | 13-14 | 11 | 12 | 11 | 13 | 11 | 30 | 18 | 9-9 | 12 | 11 | 23 | 16 | 21 | 27 | 13-13-14-14 | 10 | 11 | 20-20 | 16 | 13 | 18 | 19 | 34-38 | 11 | 10 | 12 | 8 | 15-16 | 8 | 11 | 10 | 8 | 11 | 11 | 14 | 21-24 | 15 | 11 | 12 | 12 | 15 | 8 | 14 | 21 | 24 | 17 | 13 | 11 | 13 | 10 | 11 | 11 | 12 | 31 | 15 | 8 | 16 | 11 | 21 | 27 | 22 | 11 | 11 | 12 | 13 | 11 | 9 | 9 | 11 | 10 | 10 | 12 | 27 | 10 | 12 | 21 | 13 | 11 | 11 | 28 | 15 | 17 | 14 | 24 | 17 | 13 | 15 | 27 | 12 | 21 | 19 | 11 | 14 | 19 | 9 | 11 | 11 |
| 172686 | K Shapsug | Russian Federation | G-U1 | 14 | 23 | 15 | 10 | 13-14 | 11 | 12 | 11 | 12 | 11 | 30 | 16 | 9-9 | 11 | 11 | 23 | 16 | 20 | 29 | 13-13-13-13 | 10 | 11 | 20-20 | 17 | 13 | 16 | 16 | 33-36 | 11 | 10 | 11 | 8 | 15-16 | 8 | 11 | 10 | 8 | 11 | 11 | 14 | 21-22 | 15 | 10 | 12 | 12 | 15 | 8 | 13 | 21 | 25 | 18 | 13 | 11 | 13 | 10 | 11 | 11 | 12 | 31 | 15 | 8 | 16 | 12 | 21 | 27 | 21 | 11 | 11 | 12 | 13 | 11 | 9 | 9 | 12 | 10 | 10 | 12 | 27 | 10 | 12 | 22 | 14 | 11 | 11 | 22 | 15 | 18 | 14 | 24 | 19 | 13 | 15 | 27 | 12 | 21 | 18 | 11 | 14 | 19 | 9 | 12 | 11 |
| TMRCA - Time to Most Recent Common Ancestor / Расстояние до ближайшего общего предка (мутация 0.004..0.0075 FTDNA; поколение 24-30 лет) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1176 ± 300 лет | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 172686 | K Shapsug | Russian Federation | G-U1 | 14 | 23 | 15 | 10 | 13-14 | 11 | 12 | 11 | 12 | 11 | 30 | 16 | 9-9 | 11 | 11 | 23 | 16 | 20 | 29 | 13-13-13-13-13-14-16 | 10 | 11 | 20-20 | 17 | 13 | 16 | 16 | 33-36 | 11 | 10 | 11 | 8 | 15-16 | 8 | 11 | 10 | 8 | 11 | 11 | 14 | 21-22 | 15 | 10 | 12 | 12 | 15 | 8 | 13 | 21 | 25 | 18 | 13 | 11 | 13 | 10 | 11 | 11 | 12 | 31 | 15 | 8 | 16 | 12 | 21 | 27 | 21 | 11 | 11 | 12 | 13 | 11 | 9 | 9 | 12 | 10 | 10 | 12 | 27 | 10 | 12 | 22 | 14 | 11 | 11 | 22 | 15 | 18 | 14 | 24 | 19 | 13 | 15 | 27 | 12 | 21 | 18 | 11 | 14 | 19 | 9 | 12 | 11 |
| 163132 | Mustafa Cerkesoglu (Husht) Circassia Shapsug, Trabzon | Turkey | G-M201 | 14 | 23 | 15 | 10 | 13-14 | 11 | 12 | 11 | 13 | 11 | 31 | 17 | 9-9 | 11 | 11 | 23 | 16 | 20 | 29 | 13-13-14-14 | 11 | 11 | 20-20 | 16 | 13 | 16 | 20 | 35-36 | 11 | 10 | 11 | 8 | 15-16 | 8 | 12 | 10 | 8 | 12 | 11 | 14 | 21-22 | 17 | 10 | 12 | 13 | 14 | 8 | 12 | 21 | 24 | 17 | 13 | 11 | 14 | 10 | 11 | 11 | 11 | 35 | 15 | 8 | 17 | 12 | 21 | 27 | 21 | 11 | 10 | 12 | 13 | 11 | 9 | 9 | 11 | 10 | 10 | 12 | 27 | 10 | 12 | 21 | 14 | 11 | 13 | 29 | 15 | 18 | 16 | 24 | 19 | 13 | 15 | 27 | 12 | 22 | 18 | 11 | 14 | 17 | 9 | 11 | 11 |
| TMRCA - Time to Most Recent Common Ancestor / Расстояние до ближайшего общего предка (мутация 0.004..0.0075 FTDNA; поколение 24-30 лет) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1104 ± 276 лет | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 299046 | Khuta Gulordava, b. c. 1770, Kitsia, Samegrelo | Georgia | G-M201 | 14 | 22 | 15 | 10 | 16-16 | 11 | 12 | 11 | 13 | 10 | 30 | 17 | 9-9 | 11 | 12 | 24 | 16 | 21 | 28 | 13-13-14-14 | 10 | 10 | 20-21 | 16 | 15 | 15 | 18 | 36-37 | 11 | 10 | 11 | 8 | 15-16 | 8 | 11 | 11 | 8 | 11 | 10 | 12 | 20-21 | 15 | 10 | 12 | 12 | 16 | 8 | 13 | 21 | 22 | 16 | 13 | 11 | 13 | 10 | 11 | 11 | 13 | 30 | 16 | 8 | 16 | 11 | 23 | 28 | 20 | 9 | 11 | 13 | 12 | 12 | 9 | 10 | 11 | 10 | 11 | 12 | 28 | 10 | 12 | 22 | 14 | 9 | 10 | 24 | 15 | 17 | 12 | 23 | 18 | 12 | 16 | 27 | 12 | 21 | 18 | 11 | 14 | 17 | 9 | 12 | 11 |
| 283822 | Almaskhan Buzaladze Born 1870 | Georgia | G-P303 | 14 | 22 | 15 | 11 | 13-14 | 11 | 12 | 11 | 13 | 11 | 30 | 18 | 9-9 | 12 | 11 | 23 | 16 | 21 | 27 | 13-13-14-14 | 10 | 11 | 20-20 | 16 | 13 | 18 | 19 | 34-38 | 11 | 10 | 12 | 8 | 15-16 | 8 | 11 | 10 | 8 | 11 | 11 | 14 | 21-24 | 15 | 11 | 12 | 12 | 15 | 8 | 14 | 21 | 24 | 17 | 13 | 11 | 13 | 10 | 11 | 11 | 12 | 31 | 15 | 8 | 16 | 11 | 21 | 27 | 22 | 11 | 11 | 12 | 13 | 11 | 9 | 9 | 11 | 10 | 10 | 12 | 27 | 10 | 12 | 21 | 13 | 11 | 11 | 28 | 15 | 17 | 14 | 24 | 17 | 13 | 15 | 27 | 12 | 21 | 19 | 11 | 14 | 19 | 9 | 11 | 11 |
| TMRCA - Time to Most Recent Common Ancestor / Расстояние до ближайшего общего предка (мутация 0.004..0.0075 FTDNA; поколение 24-30 лет) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2256 ± 564 года | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Kit Number |
Paternal Ancestor Name |
Country |
Haplogroup |
DYS393 |
DYS390 |
DYS19 |
DYS391 |
DYS385 |
DYS426 |
DYS388 |
DYS439 |
DYS389i |
DYS392 |
DYS389ii |
DYS458 |
DYS459 |
DYS455 |
DYS454 |
DYS447 |
DYS437 |
DYS448 |
DYS449 |
DYS464 |
DYS460 |
Y-GATA-H4 |
YCAII |
DYS456 |
DYS607 |
DYS576 |
DYS570 |
CDY |
DYS442 |
DYS438 |
DYS531 |
DYS578 |
DYF395S1 |
DYS590 |
DYS537 |
DYS641 |
DYS472 |
DYF406S1 |
DYS511 |
DYS425 |
DYS413 |
DYS557 |
DYS594 |
DYS436 |
DYS490 |
DYS534 |
DYS450 |
DYS444 |
DYS481 |
DYS520 |
DYS446 |
DYS617 |
DYS568 |
DYS487 |
DYS572 |
DYS640 |
DYS492 |
DYS565 |
DYS710 |
DYS485 |
DYS632 |
DYS495 |
DYS540 |
DYS714 |
DYS716 |
DYS717 |
DYS505 |
DYS556 |
DYS549 |
DYS589 |
DYS522 |
DYS494 |
DYS533 |
DYS636 |
DYS575 |
DYS638 |
DYS462 |
DYS452 |
DYS445 |
Y-GATA-A10 |
DYS463 |
DYS441 |
Y-GGAAT-1B07 |
DYS525 |
DYS712 |
DYS593 |
DYS650 |
DYS532 |
DYS715 |
DYS504 |
DYS513 |
DYS561 |
DYS552 |
DYS726 |
DYS635 |
DYS587 |
DYS643 |
DYS497 |
DYS510 |
DYS434 |
DYS461 |
DYS435 |
| 163132 | Mustafa Cerkesoglu (Husht) Circassia Shapsug, Trabzon | Turkey | G-M201 | 14 | 23 | 15 | 10 | 13-14 | 11 | 12 | 11 | 13 | 11 | 31 | 17 | 9-9 | 11 | 11 | 23 | 16 | 20 | 29 | 13-13-14-14 | 11 | 11 | 20-20 | 16 | 13 | 16 | 20 | 35-36 | 11 | 10 | 11 | 8 | 15-16 | 8 | 12 | 10 | 8 | 12 | 11 | 14 | 21-22 | 17 | 10 | 12 | 13 | 14 | 8 | 12 | 21 | 24 | 17 | 13 | 11 | 14 | 10 | 11 | 11 | 11 | 35 | 15 | 8 | 17 | 12 | 21 | 27 | 21 | 11 | 10 | 12 | 13 | 11 | 9 | 9 | 11 | 10 | 10 | 12 | 27 | 10 | 12 | 21 | 14 | 11 | 13 | 29 | 15 | 18 | 16 | 24 | 19 | 13 | 15 | 27 | 12 | 22 | 18 | 11 | 14 | 17 | 9 | 11 | 11 |
| 218868 | Vardan Dadiani, b.c. XII c. | Georgia | G-M201 | 13 | 22 | 15 | 10 | 15-18 | 11 | 12 | 12 | 11 | 10 | 29 | 16 | 9-9 | 11 | 11 | 26 | 16 | 21 | 28 | 13-13-14-14 | 10 | 10 | 19-19 | 15 | 15 | 16 | 18 | 35-35 | 12 | 10 | 11 | 8 | 15-16 | 8 | 11 | 10 | 8 | 11 | 10 | 12 | 21-22 | 14 | 10 | 12 | 12 | 14 | 8 | 12 | 21 | 22 | 16 | 13 | 11 | 13 | 10 | 11 | 11 | 12 | ||||||||||||||||||||||||||||||||||||||||||||
| TMRCA - Time to Most Recent Common Ancestor / Расстояние до ближайшего общего предка (мутация 0.004..0.0075 FTDNA; поколение 24-30 лет) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1800 ± 450 лет | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 306115 | Chaguna Karbaia, b. c. 1775, Darcheli, Samegrelo | Georgia | G-M201 | 14 | 23 | 15 | 10 | 13-13 | 11 | 12 | 11 | 12 | 11 | 28 | 18 | 9-9 | 11 | 11 | 23 | 16 | 20 | 31 | 13-13-13-14 | 10 | 11 | 20-21 | 17 | 12 | 15 | 18 | 35-36 | 12 | 10 | 11 | 8 | 15-16 | 8 | 11 | 10 | 8 | 12 | 11 | 14 | 21-22 | 15 | 10 | 12 | 12 | 13 | 8 | 12 | 21 | 23 | 19 | 13 | 11 | 13 | 10 | 11 | 11 | 12 | 29 | 15 | 8 | 16 | 12 | 21 | 27 | 21 | 11 | 11 | 11 | 13 | 11 | 9 | 9 | 11 | 10 | 10 | 12 | 27 | 10 | 11 | 21 | 14 | 11 | 13 | 26 | 15 | 19 | 14 | 24 | 18 | 13 | 15 | 27 | 12 | 21 | 18 | 11 | 14 | 16 | 9 | 12 | 11 |
| 243083 | Akoev, Ak'oti, North Ossetia | Russian Federation | G-M201 | 13 | 23 | 15 | 10 | 13-14 | 11 | 12 | 11 | 12 | 11 | 29 | 16 | 9-9 | 11 | 11 | 23 | 16 | 20 | 28 | 13-13-13-13 | 11 | 12 | 20-20 | 16 | 13 | 16 | 16 | 34-35 | 11 | 10 | 11 | 8 | 16-16 | 8 | 11 | 10 | 8 | 13 | 11 | 14 | 21-22 | 16 | 10 | 12 | 12 | 15 | 8 | 13 | 21 | 26 | 19 | 13 | 11 | 13 | 10 | 11 | 11 | 12 | 31 | 15 | 8 | 16 | 12 | 22 | 27 | 21 | 11 | 11 | 13 | 12 | 11 | 9 | 9 | 12 | 10 | 10 | 11 | 28 | 10 | 12 | 21 | 14 | 11 | 11 | 20 | 15 | 18 | 17 | 24 | 18 | 13 | 15 | 27 | 12 | 21 | 18 | 11 | 14 | 18 | 9 | 12 | 11 |
| TMRCA - Time to Most Recent Common Ancestor / Расстояние до ближайшего общего предка (мутация 0.004..0.0075 FTDNA; поколение 24-30 лет) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1320 ± 330 лет | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 216684 | Kaikhosro Abashidze, b.c. 17??, Georgia | Georgia | G-M201 | 13 | 23 | 15 | 10 | 15-16 | 11 | 12 | 10 | 12 | 10 | 28 | 18 | 9-9 | 11 | 11 | 24 | 16 | 21 | 29 | 13-14-14-14 | 10 | 10 | 21-21 | 14 | 15 | 14 | 18 | 34-37 | 12 | 10 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 302135 | Kharchilava Vasil, b. 1878 and d. 1977 | Georgia | G-M201 | 14 | 22 | 15 | 10 | 16-17 | 11 | 12 | 12 | 12 | 10 | 28 | 16 | 9-9 | 11 | 12 | 24 | 16 | 21 | 29 | 13-14-14-14 | 10 | 10 | 19-20 | 15 | 15 | 16 | 20 | 35-39 | 12 | 10 | 11 | 8 | 15-16 | 8 | 11 | 10 | 8 | 12 | 10 | 12 | 21-22 | 14 | 10 | 12 | 12 | 15 | 8 | 13 | 21 | 21 | 16 | 13 | 11 | 13 | 10 | 11 | 11 | 12 | ||||||||||||||||||||||||||||||||||||||||||||
| TMRCA - Time to Most Recent Common Ancestor / Расстояние до ближайшего общего предка (мутация 0.004..0.0075 FTDNA; поколение 24-30 лет) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1008 ± 252 года | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 172686 | K Shapsug | Russian Federation | G-U1 | 14 | 23 | 15 | 10 | 13-14 | 11 | 12 | 11 | 12 | 11 | 30 | 16 | 9-9 | 11 | 11 | 23 | 16 | 20 | 29 | 13-13-13-13-13-14-16 | 10 | 11 | 20-20 | 17 | 13 | 16 | 16 | 33-36 | 11 | 10 | 11 | 8 | 15-16 | 8 | 11 | 10 | 8 | 11 | 11 | 14 | 21-22 | 15 | 10 | 12 | 12 | 15 | 8 | 13 | 21 | 25 | 18 | 13 | 11 | 13 | 10 | 11 | 11 | 12 | 31 | 15 | 8 | 16 | 12 | 21 | 27 | 21 | 11 | 11 | 12 | 13 | 11 | 9 | 9 | 12 | 10 | 10 | 12 | 27 | 10 | 12 | 22 | 14 | 11 | 11 | 22 | 15 | 18 | 14 | 24 | 19 | 13 | 15 | 27 | 12 | 21 | 18 | 11 | 14 | 19 | 9 | 12 | 11 |
| 259023 | Laz | Georgia | G-M201 | 14 | 23 | 15 | 10 | 13-14 | 12 | 12 | 11 | 12 | 12 | 29 | 17 | 9-9 | 11 | 11 | 24 | 15 | 20 | 29 | 13-13-13-14 | 11 | 11 | 20-20 | 17 | 13 | 15 | 17 | 34-37 | 11 | 10 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TMRCA - Time to Most Recent Common Ancestor / Расстояние до ближайшего общего предка (мутация 0.004..0.0075 FTDNA; поколение 24-30 лет) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 912 ± 228 лет | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The conclusion on TMRCA:
The averaged Time to a most recent common male ancestor on G-M201 for the listed in Table 1: 1551 ± 332 years.
- The main Western Caucasus Y-DNA haplogroup "G"
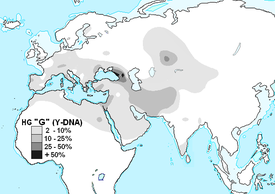
Western Caucasian (Circassians, Abkhazians, Megrelians, Georgians, Ossetians) Y-chromosome haplogroups conclusions
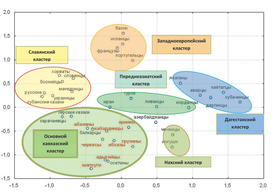
1. The main share of the Y-chromosome in the Western Caucasus falls on haplogroups G2-P287 (44,1%) and J2-M172 (20,1%), which indicates two main common ancestors in the male line.
2. Analysis of the Y-chromosomes of Abkhazians showed results similar to the previous conclusion: haplogroup G2a >50% (G2a1a: 12%, G2a*: 24%, G2a3b1 (G2a1c2a): 21%) and haplogroup J2a ~20% (J2*: 5%, J2a4b*: 7%, J2a4b1: 2% [according to Balanovsky]; J2a*: 14.8%, J2a2*: 8.6%, J2a2a: 2.5%, J2b: 0.6% [according to Litvinov]).
3. The separation of Mingrelians and Abkhazians shows an almost identical picture of the mixture of all Y-chromosome globular groups.
4. The proximity of the studied populations to each other determines the presence of similar haplogroups. For example, in Eastern Georgia, there is a border influence of neighbors, which is expressed in a greater number of carriers of haplogroups R1b and J1.
5. A common ancestor G-P303 lived approximately 11500 years ago possibly in Levant.
6. A common ancestor of G-P303 for the entire Caucasus lived approximately 5,345 years ago.
7. A common ancestor G-P303 for Abkhazians, Shapsugs and Circassians lived approximately 2445 years ago.
8. Apparently, haplogroup G2a came to the Caucasus from Europe, where it is found in ancient burials dating back 5,000 years. It looks like carriers of the haplogroup G2a passed from Europe through Asia Minor to Mesopotamia, and from there they ascended to the Caucasus and Northern China.
9. Average distance to the closest common male ancestor according to G-M201 for the species listed in Table 1: 1551 ± 332 years.
10. Among the Abkhaz-Adyghe peoples, the major haplogroups are G2a3b-P303, G2a1a-P18, J2-M172 and R1a1-M198, among the Armenian populations – haplogroups G1-M285 and J2a2-M67, among the Georgians – G2a3b-P303, G2a1a-P18 and J2-M172. The peoples of the Western Caucasus and Transcaucasia in the genetic space of Eurasia form a single cluster, isolated from other gene pools of Eurasia. The revealed predominance of the Near Asian substrate (G2a3b-P303, G2a1a-P18 and J2-M172) indicates their common origin. The high frequency of the Eurasian haplogroup R1a1-M198 and the low frequency of the subbranch R1a1a1g-M458 characteristic of Europe among the Abkhaz-Adyghe peoples is associated with the migration of peoples from the Eurasian steppe, and not with the expansion of the Slavic peoples.